- CDFA Home
- Plant Health
- PPD
- Entomology
- Scenopinidae

Plant Pest Diagnostics Center - Entomology Laboratory
Research Programs - Insecta: Diptera Asiloidea: Scenopinidae
Window Flies of the World (Diptera: Scenopinidae)

Drs. Shaun Winterton & Steve Gaimari
Welcome to the Window-fly website. The study of scenopinid evolution and biodiversity is an important part of the Diptera research programs of Steve Gaimari and Shaun Winterton, who both work on Asiloidea and 'Therevoid'-clade systematics. The current project is supported by an NSF grant to both of them (DEB-0614213). Scenopinidae are a poorly studied group of flies that show a tremendous wealth of unexplored morphological and biological diversity. This site is still under construction, but already contains an extensive bibliogrpahy of Scenopinidae literature, a checklist of world species, and a dichotomous key to genera. Shortly, a full catalogue of the family will be finished, and interactive Lucid3 keys, fact sheets and image galleries will be placed on this site. Please check back frequently for updates.
Scenopinidae, the window-flies, are a cosmopolitan group of flies with adults that are typically small and dark with a body size rarely greater than 5.0 mm. While found in a variety of habitats, by far the greatest diversity of this group is in arid regions, where the dry sandy soils provide a suitable habitat for the larvae. As larvae, scenopinids are elongate, fossorial predators of arthropods in friable soils and leaf litter, although larvae have also been reared from, or suspected of breeding in, habitats as diverse as galleries of wood-boring insects, birds nests, mammal nest holes, termite mounds, bat guano, beehives and stored products (Kelsey 1969, Rahman et al. 1981, Yucel 1988, Gnaspini 1989, Yao & Lo 1992, Dobson 1999). A single case of human urogenital myasis is recorded for a larva of Scenopinus Latreille (Thompson et al. 1970), but this appears to be exceptional.
Some adult scenopinids apparently do not to feed (e.g. Belosta Hardy), but many are nectar and honeydew feeders and are often collected by sweeping flowers and foliage (Kelsey 1975a, 1987). Adults of some species (e.g. Scenopinus spp.) are often collected on windows inside human dwellings due to their predation as larvae on pests associated with human activities; hence leading to their common name as window-flies (Kelsey 1969, Kelsey 1981a,b,c, Cole 1923, Melander 1950).
Scenopinidae are placed in the superfamily Asiloidea, and had long been accepted as the sister group to the stiletto-flies (Therevidae) based on secondary segmentation of the larval abdomen (Woodley 1989). Using morphological evidence, Sinclair et al. (1994) and Yeates (1994, 2002) showed that Scenopinidae and Therevidae are closely related to Apsilocephalidae, a hypothesis supported by 28S ribosomal and elongation factor-1 alpha (EF1-a) sequence data (Yang et al. 2000, Wiegmann et al. 2003) Using 28S rDNA Yeates et al. (2003) placed the newly described family Ocoidae as sister to Scenopinidae and Therevidae. While the monophyly of the 'therevoid' clade is relatively well supported in their analysis, the relationships between the four individual families are not. Unfortunately the larvae of both Apsilocephalidae and Ocoidae are unknown, so we cannot determine whether they too have secondarily segmented abdomens. Moreover, several authors have suggested that the lack of clear synapomorphies supporting the Therevidae may be indicative of Therevidae being paraphyletic with respect to Scenopinidae (Woodley 1989, Yeates 1992, Yeates & Wiegmann 1999).
Scenopinidae are placed in the superfamily Asiloidea, and had long been accepted as the sister group to the stiletto-flies (Therevidae) based on secondary segmentation of the larval abdomen (Woodley 1989). Using morphological evidence, Sinclair et al. (1994) and Yeates (1994, 2002) showed that Scenopinidae and Therevidae are closely related to Apsilocephalidae, a hypothesis supported by 28S ribosomal and elongation factor-1 alpha (EF1-a) sequence data (Yang et al. 2000, Wiegmann et al. 2003) Using 28S rDNA Yeates et al. (2003) placed the newly described family Ocoidae as sister to Scenopinidae and Therevidae. While the monophyly of the 'therevoid' clade is relatively well supported in their analysis, the relationships between the four individual families are not. Unfortunately the larvae of both Apsilocephalidae and Ocoidae are unknown, so we cannot determine whether they too have secondarily segmented abdomens. Moreover, several authors have suggested that the lack of clear synapomorphies supporting the Therevidae may be indicative of Therevidae being paraphyletic with respect to Scenopinidae (Woodley 1989, Yeates 1992, Yeates & Wiegmann 1999).
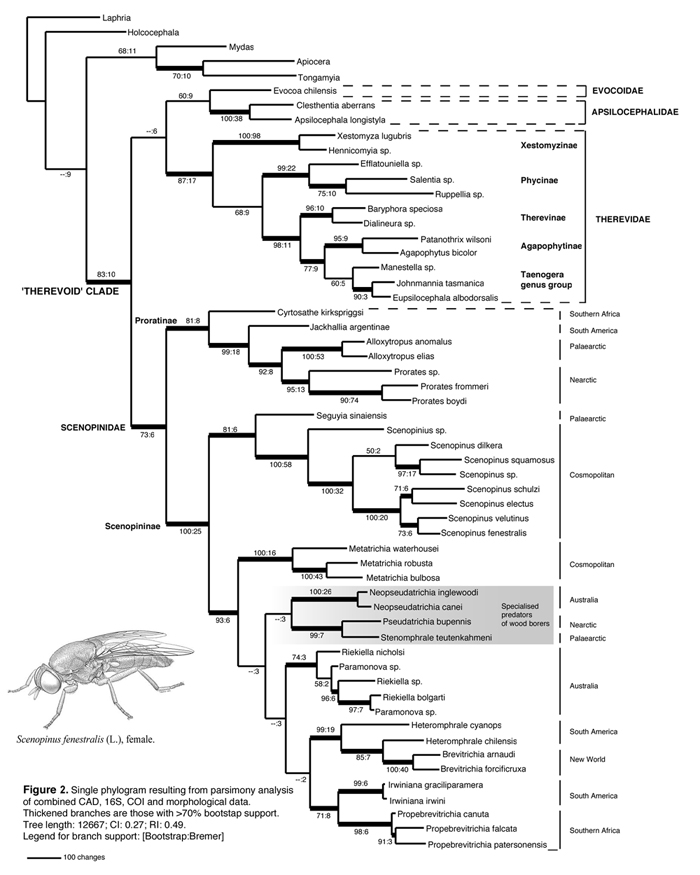
Twenty-four extant genera of Scenopinidae are presently recognised; (Table 1) containing more than 420 described species worldwide (Kelsey 1973b, Yeates 1992, Nagatomi et al. 1994). Kelsey (1969 to 1989) contributed significantly to our present knowledge of Scenopinidae taxonomy, describing numerous new species and genera from all major biogeographical regions. Unfortunately, this work was not based on rigorous cladistic methodology, so the predictive value of Kelsey's work is limited. The genus and tribal level relationships of the Scenopinidae are not known and discovering these is the next step in furthering our understanding of the group. To date, only Yeates (1992) and Nagatomi et al. (1994) have examined the Scenopinidae in a phylogenetic context. In a cladistic analysis of 25 adult morphological characters, Yeates (1992) justified the inclusion of Prorates, Caenotus, Alloxytropus and Caenotoides in Scenopinidae and erected the first robust subfamilial classification of the family. Three subfamilies are now recognized, the basal Caenotinae (containing Caenotus), the Proratinae (containing Prorates, Alloxytropus, Caenotoides, Jackhallia Nagatomi & Lui and Acaenotus Nagatomi & Yanagida) and the Scenopininae (containing all other scenopinid genera). Unfortunately, Yeates (1992) did not examine scenopinine relationships and included only two genera from the subfamily in the analysis. Nagatomi et al. (1994) subsequently examined the Proratinae (inclusive of Caenotus), describing two new genera (Acaenotus and Jackhallia) and proposing several competing hypotheses of generic relationships within the subfamily. Such contradictory proposals by Nagatomi et al. (1994) has not generally lead to better understanding of proratine relationships, but did provide important detailed morphological studies of the group. Recently Winterton & Metz (2005) described an entirely new genus of scenopinid from Namibia (i.e. Cyrtosathe Winterton & Metz) that exhibits characteristics from all three subfamilies. Cyrtosathe is presently placed as incertae sedis within Scenopinidae until a complete phylogenetic analysis and detailed reappraisal of the classification can be undertaken to place this new taxon. Just when we thought that the relationships of Scenopinidae subfamilies had started to become clearer, the discovery of this enigmatic new genus has forced us to reevaluate our basic definitions of the subfamilies themselves. Moreover, the phylogenetic relationships of the bulk of Scenopinidae, i.e. those genera placed in the Scenopininae and representing over 85% of the species diversity of the family, are still unknown and remain a significant challenge. Clearly, the relationships of the 'basal' scenopinid subfamilies (Caenotinae and Proratinae) with respect to the Scenopininae, along with scenopinine relationships, need to be tested fully with more extensive exemplar sampling from all groups. These are central questions for my research on these flies.
Family: SCENOPINIDAE
- Caenotinaeod
- hospes Melander 1950: 149, Nearctic
- inornatus Cole 1923a: 16, Nearctic
- mexicanus Nagatomi & Yanagida 1994: 159, Nearctic
- minutus Cole 1923a: 15, Nearctic
- tanyrhynchus Metz 2003 : 4, Nearctic
- Proratinae
- canus (Melander) 1950: 149 (Caenotus), Nearctic
- anomalus Bezzi 1925b: 187, Palearctic
- bezzi Paramonov 1929: 83, Palearctic
- elias Winterton & Kerr 2006 : 42, Palearctic
- kerzhneri Zaitzev 1972b: 860, Palearctic
- lehri Zaitzev 1972: 455, Palearctic
- californica Hall 1972: 48, Nearctic
- idahoensis Hall 1972: 49, Nearctic
- mexicana Hall 1972: 49, Nearctic
- argentinae Nagatomi & Liu 1994: 167, Neotropical
- arctos Hall 1972: 42, Nearctic
- ballmeri Nagatomi & Liu 1994: 171, Nearctic
- boydi Hall 1972: 41, Nearctic
- claripennis Melander 1906: 373, Nearctic
- frommeri Hall 1972: 45, Nearctic
- melanderi Hall 1972: 43, Nearctic
- nigrescens Hall 1972: 44, Nearctic
- painteri Nagatomi & Liu 1994: 181, Nearctic
- simplex Grimaldi & Cumming 1999: 30, Amber, New Jersey (Mesozoic) [extinct]
- Scenopininae
- albicomula Kelsey 1971d: 34, Nearctic
- albipilosa D.E. Hardy 1944a: 38, Nearctic
- flaviceps (Coquillett) 1902: 102 (Pseudatrichia), Nearctic
- pilosa (Coquillett) 1902: 102 (Pseudatrichia), Nearctic
- scutulamacula Kelsey 1971d: 35, Nearctic
- telfordi Kelsey 1969: 278, Nearctic
- termitophaga Kelsey 1969: 278, Nearctic
- viticolapennis Kelsey 1969: 281, Nearctic
- addacifrons Kelsey 1969: 169, Nearctic
- albanota Kelsey 1971d: 17, Nearctic
- arena Kelsey 1971d: 18, Nearctic
- argentaurba Kelsey 1971d: 19, Nearctic
- arnaudi Kelsey 1969: 171, Nearctic
- aspinosa Kelsey 1969: 172, Nearctic
- badiclitella Kelsey 1971d: 19, Nearctic
- beameri Kelsey 1969: 174, Nearctic
- [=griseola Cresson (nec Coquillett) 1907: 113]
- boharti Kelsey 1969: 175, Nearctic
- castanea Kelsey 1969: 177, Nearctic
- coquilletti Kelsey 1969: 177, Nearctic
- [=griseola Coquillett 1900: 501 ( part)]
- daileyi Kelsey 1971d: 20, Nearctic
- davisi Kelsey 1969: 180, Nearctic
- dicksoni Kelsey 1969: 182, Nearctic
- downeyi Kelsey 1969: 184, Nearctic
- flocki Kelsey 1969: 186, Nearctic
- forficicruxa Kelsey 1971d: 21, Nearctic
- griffini Kelsey 1969: 186, Nearctic
- griseola (Coquillett) 1900: 501 (Pseudatrichia), Nearctic
- halli Kelsey 1969: 192, Nearctic
- helenae (James) 1938: 22 (Pseudatrichia), Nearctic
- hodgdeni Kelsey 1969: 194, Nearctic
- inferacanna Kelsey 1971d: 21, Nearctic
- insulana (Cole) 1923b: 462 (Pseudatrichia), Nearctic
- irwini Kelsey 1971d: 22, Nearctic
- kerni Kelsey 1969: 196, Nearctic
- melanderi Kelsey 1969: 199, Nearctic
- minuta Kelsey 1969: 201, Nearctic
- miraloma Kelsey 1971d: 23, Nearctic
- nayariti Kelsey 1971d: 23, Nearctic
- nevada Kelsey 1971d: 24, Nearctic
- oculivirida Kelsey 1969: 202, Nearctic
- ordwayi Kelsey 1969: 204, Nearctic
- palida Kelsey 1971d: 25, Nearctic
- palmacana Kelsey 1971d: 26, Nearctic
- piscifonta Kelsey 1971d: 27, Nearctic
- powelli Kelsey 1974b: 139, Nearctic
- pruinosa Kelsey 1969: 206, Nearctic
- salvadorensis Kelsey 1974b: 139, Nearctic
- schlingeri Kelsey 1969: 207, Nearctic
- scitulaesca Kelsey 1969: 209, Nearctic
- septispina Kelsey 1971d: 28, Nearctic
- timberlakei Kelsey 1969: 210, Nearctic
- wilcoxi Kelsey 1971d: 29, Nearctic
- yucatani Kelsey 1969: 212, Nearctic
- nigra Kelsey 1969: 163, Palearctic
- robusta Kröber 1923: 75, Palearctic
- chilensis (Kröber) 1928: 31 (Pseudatrichia), Neotropical
- cyanops (Edwards) 1932: 259 (Pseudomphrale), Neotropical
- glabrifrons Kelsey 1971b: 282, Neotropical
- graciliparamera Kelsey 1971b: 283, Neotropical
- irwini Kelsey 1971b: 281, Neotropical
- asiatica Krivosheina & Krivosheina 1999: 849, Palearctic
- bilituua Kelsey 1981: 6, Palearctic
- bulbosus (Osten Sacken) 1877: 275 (Scenopinus), Nearctic
- deserticola Krivosheina & Krivosheina 1999: 851, Palearctic
- friedbergi Krivosheina & Krivosheina 1999: 852, Palearctic
- lophyrosoma (Speiser) 1920: 216 (Pseudomphrale), Afrotropical
- mongolica Kelsey 1981: 85, Palearctic
- nigeriana Kelsey 1984: 57, Afrotropical
- palaestinensis (Kröber) 1937: 229 (Pseudomphrale), Palearctic
- papuana Kelsey 1970a: 44, Australasian
- pria Yeates & Grimaldi 1993: 2, Amber, Dominican Republic (Oligo-Miocene) [extinct]
- robusta Kröber 1913: 180, Neotropical
- stevensoni (Bezzi) 1925: 96 (Pseudomphrale), Afrotropical
- [=africana Lindner, 1962: 2]
- thailandica Kelsey 1970a: 46, Oriental
- waterhousei (Paramonov) 1955: 640 (Pseudomphrale), Australasian
- canei Kelsey 1969: 300, Australasian
- [=mariaensis Paramonov (nec. G.H. Hardy) 1955: 639]
- inglewoodi Kelsey 1970b: 141, Australasian
- kewi Kelsey 1969: 300, Australasian
- mariaensis (G.H. Hardy) 1933: 419 (Pseudatrichia), Australasian
- monteithi Kelsey 1970b: 144, Australasian
- arila Kelsey 1987:10, Australasian
- brevipalpa Kelsey 1970b: 127, Australasian
- bungabandii Kelsey 1975a: 205, Australasian
- burrijuanna Kelsey 1987:11, Australasian
- bushi Kelsey 1971c: 203, Australasian
- collessi (Kelsey) 1969: 307 (Riekiella), Australasian
- complexa (Kelsey) 1969: 309 (Riekiella), Australasian
- dycei (Kelsey) 1969: 310 (Riekiella), Australasian
- emarginata Kelsey 1970b: 129, Australasian
- fonsamniculi Kelsey 1980a: 9, Australasian
- hamata Kelsey 1980a: 10, Australasian
- mangarra Kelsey 1987: 12, Australasian
- poocheraiana Kelsey 1987: 13, Australasian
- quadrifida (Kelsey) 1969: 313 (Riekiella), Australasian
- taylori (Kelsey) 1969: 315 (Riekiella), Australasian
- uncata (Kelsey) 1969: 315 (Riekiella), Australasian
- zentae Kelsey 1975a: 207, Australasian
- lobosa Kelsey 1969: 320, Australasian
- spicata Kelsey 1975a: 208, Australasian
- westralica (Paramonov) 1955: 641 (Pseudomphrale), Australasian
- lobosa Kelsey 1969: 320, Australasian
- spicata Kelsey 1975a: 208, Australasian
- westralica (Paramonov) 1955: 641 (Pseudomphrale), Australasian
- kelseyi Krivosheina 1980: 198, Palearctic
- mateui Kelsey 1969: 289, Palearctic, Afrotropical
- stenogaster (Séguy) 1931: 114 (Scenopinus), Afrotropical
- violacea Kelsey 1969: 291, Afrotropical
- bonnieana Kelsey 1976: 961, Afrotropical
- botterkloofensis Kelsey 1976: 963, Afrotropical
- canuta Kelsey 1976: 963, Afrotropical
- dionaeforma Kelsey 1976: 964, Afrotropical
- falcata Kelsey 1976: 967, Afrotropical
- gobabebensis Kelsey 1976: 968, Afrotropical
- irwini Kelsey 1976: 970, Afrotropical
- langebaanensis Kelsey 1976: 971, Afrotropical
- pakhiusensis Kelsey 1976: 973, Afrotropical
- patersonensis Kelsey 1976: 975, Afrotropical
- quadrispina Kelsey 1971a: 170, Afrotropical
- serowensis Winterton 2005 : 2, Afrotropical
- stuckenbergi Kelsey 1969: 283, Afrotropical
- swakopensis Kelsey 1976: 976, Afrotropical
- turneri Kelsey 1969: 284, Afrotropical
- albocincta Van Duzee 1926: 164, Nearctic
- atombomba Kelsey 1969: 225, Nearctic
- [=unicolor Cresson (not Coquillett), 1907: 113. (part)]
- bakeri Kelsey 1969: 227, Nearctic
- barnesi Kelsey 1969: 229, Nearctic
- biacristerna Kelsey 1969: 230, Nearctic
- bryanti Kelsey 1969: 231, Nearctic
- bupennis Kelsey 1969: 232, Nearctic
- cajoni Kelsey 1969: 234, Nearctic
- cloudcrofti Kelsey 1969: 236, Neartic
- [=unicolor Cresson (nec. Coquillett) 1907: 113. (part)]
- cockerelli Kelsey 1969: 236, Nearctic
- concava Kelsey 1969: 239, Nearctic
- convexa Kelsey 1969: 241, Nearctic
- eaithales Kelsey 1969: 241, Nearctic
- evergreeni Kelsey 1969: 244, Nearctic
- garretti Kelsey 1969: 244, Nearctic
- gracilipennis Kelsey 1969: 247, Nearctic
- granti Kelsey 1974b: 137, Nearctic
- howdeni Kelsey 1969: 247, Nearctic
- idahoensis Kelsey 1971d: 30, Nearctic
- jamesi Kelsey 1969: 250, Nearctic
- leechi Kelsey 1969: 250, Nearctic
- longiventris Kelsey 1969: 253, Neotropical
- longurio (Loew) 1866: 42 (Atrichia), Nearctic
- macalpeni Kelsey 1974b: 137, Nearctic
- melanderi Kelsey 1969: 256, Nearctic
- morlani Kelsey 1969: 258, Nearctic
- nordeni Kelsey 1969: 260, Nearctic
- parva D.E. Hardy 1944a: 36, Nearctic
- parvipennis Kelsey 1969: 260, Nearctic
- peromysci Kelsey 1969: 264, Nearctic
- punctulata D.E. Hardy 1944: 104, Neotropical
- rufitruncula Kelsey 1971d: 31, Nearctic
- sabroskyi Kelsey 1969: 266, Nearctic
- saccharcupa Kelsey 1969: 268, Nearctic
- schlingeri Kelsey 1971d: 32, Nearctic
- tomichi Kelsey 1974b: 138, Nearctic
- toupeta Kelsey 1969: 270, Nearctic
- truncata Kelsey 1969: 270, Nearctic
- unicolor Coquillett 1900: 500, Nearctic
- clausa (Loew) 1873: 150 (Scenopinus), Palearctic
- crenata (Becker) 1913: 550 (Omphrale), Palearctic
- dubiosa (Becker) 1913: 551 (Omphrale), Palearctic
- longirostris (Becker) 1913: 549 (Omphrale), Palearctic
- miramisho Hassan & El-Hawagry 2001: 3, Palearctic
- arilaringa Kelsey 1987:16, Australasian
- arnhemi Kelsey 1975a: 210, Australasian
- bicornis Paramonov 1955: 653, Australasian
- binnaliweanda Kelsey 1987:18, Australasian
- bolgarti (Kelsey) 1969: 55 (Scenopinus), Australasian
- burririmbija Kelsey 1987:19, Australasian
- cetrata Kelsey 1971c: 196, Australasian
- cornuta Kelsey 1971c: 197, Australasian
- culbara Kelsey 1987: 20, Australasian
- cuspidata Kelsey 1971c: 198, Australasian
- denticulata Kelsey 1971c: 200, Australasian
- kilyalpa Kelsey 1987: 21, Australasian
- longitermina Kelsey 1975a: 211, Australasian
- longiventris Kelsey 1969: 312, Australasian
- malkana Kelsey 1987: 22, Australasian
- maloga Kelsey 1987: 23, Australasian
- murrilla Kelsey 1987: 24, Australasian
- nancyae Kelsey 1987: 25, Australasian
- nicholsi Kelsey 1971c: 200, Australasian
- noona Kelsey 1987: 27, Australasian
- rossi Kelsey 1970b: 123, Australasian
- rupari Kelsey 1987: 27, Australasian
- tamala Kelsey 1987: 29, Australasian
- woodwardi Kelsey 1971c: 201, Australasian
- yoota Kelsey 1987: 30, Australasian
- adventicius D.E. Hardy 1960: 329, Oceanian [velutinus group]
- adyeri Kelsey 1987: 4, Australasian [albicinctus group]
- aethiopicus (Lindner) 1955: 20 (Omphrale), Afrotropical [albicinctus group]
- afghanistanensis Kelsey 1981b: 88, Palearctic [brevicornis group]
- alalacteus Kelsey 1969: 114, Nearctic [velutinus group]
- albicinctus (Rossi) 1794: 69 (Musca), Palearctic [albicinctus group]
- [=zelleri Loew 1845: 314]
- albicomus Kelsey 1969: 17, Palearctic [fenestralis group]
- albifasciatus (D.E. Hardy) 1944a: 41 (Ompralosoma), Nearctic [velutinus group]
- albipilosus Kelsey 1971d: 7, Nearctic [velutinus group]
- angustifrons (Kröber) 1913: 197 (Omphrale), Afrotropical [albicinctus group]
- antennatus (Kröber) 1914b: 74 (Omphrale), Palearctic [fenestralis group]
- anthrax Kelsey 1969: 51, Afrotropical [albicinctus group]
- aquelonius Kelsey 1969: 20, Nearctic [fenestralis group]
- arrectus Kelsey 1975a: 198, Australasian [albicinctus group]
- ascensus Kelsey 1987: 6, Australasian [albicinctus group]
- aurantipalpus Kelsey 1971d: 7, Nearctic [velutinus group]
- balteatus Lamb 1922: 363, Afrotropical [albicinctus group]
- barberi Kelsey 1969: 115, Nearctic [velutinus group]
- beameri var. beameri (D.E. Hardy) 1944a: 43 (Omphrale), Nearctic [velutinus group]
- beameri var. fusca (D.E. Hardy) 1944a: 43 (Omphrale), Nearctic [velutinus group]
- bermudaensis Kelsey 1971: 50, Nearctic [velutinus group]
- bicuspis Kelsey 1971c: 185, Australasian [albicinctus group]
- biroi Kertész 1899: 173, Australasian [albicinctus group]
- bouvieri (Séguy) 1920: 317 (Omphrale), Afrotropical, Palearctic (introduced) [albicinctus group]
- brevicornis Loew 1873: 146, Palearctic [brevicornis group]
- breviterminus Kelsey 1969: 21, Nearctic [fenestralis group]
- brittoni Kelsey 1970b: 111, Australasian [albicinctus group]
- brunneus (Kröber) 1913: 192 (Omphrale), Afrotropical [albicinctus group]
- bryanti Kelsey 1969: 118, Nearctic [velutinus group]
- bulbapennis Kelsey 1969: 60, Palearctic [albicinctus group]
- buscki Kelsey 1969: 119, Neotropical [velutinus group]
- butleri Kelsey 1969: 121, Nearctic [velutinus group]
- canarius Kelsey 1969: 60, Palearctic [albicinctus group]
- casuarinus Kelsey 1975a: 200, Australasian [albicinctus group]
- cavifrons (Kröber) 1937: 231 (Omphrale), Nearctic [fenestralis group]
- chico Kelsey 1969: 122, Nearctic [velutinus group]
- cochisei Kelsey 1969: 124, Nearctic [velutinus group]
- cooki Kelsey 1969: 125, Nearctic [velutinus group]
- cornesi Kelsey 1984: 53, Afrotropical [brevicornis group]
- cornutus Kelsey 1981: 8, Palearctic [fenestralis group]
- cretatus Kelsey 1969: 127, Nearctic [velutinus group]
- cristafrons Kelsey 1971c: 186, Australasian [albicinctus group]
- curticornis (Kröber) 1913: 202 (Omphrale), Neotropical [velutinus group]
- curtipilosus Kelsey 1969: 63, Palearctic [albicinctus group]
- darwini Kelsey 1969: 64, Australasian [albicinctus group]
- deemingi Kelsey 1973a: 154, Afrotropical [fenestralis group]
- desertus Krivosheina 1980: 199, Palearctic [albicinctus group]
- dietricki Kelsey 1971d: 8, Nearctic [velutinus group]
- dilkera Kelsey 1987: 7, Australasian [albicinctus group]
- downesi Kelsey 1971c: 187, Australasian [albicinctus group]
- efflatouni Kelsey 1969: 24, Palearctic [fenestralis group]
- electus Adams 1904: 445, Nearctic [velutinus group]
- erugatus Kelsey 1975a: 201, Australasian [albicinctus group]
- estesi Kelsey 1971d: 3, Nearctic [fenestralis group]
- evansi Kelsey 1969: 66, Afrotropical [albicinctus group]
- eyrei Kelsey 1975a: 202, Australasian [albicinctus group]
- femoratus Macquart 1835: 7, Neotropical [velutinus group]
- fenestralis (Linnaeus) 1758: 597 (Musca), Cosmopolitan [fenestralis
group]
- [=senilis Fabricius 1794: 331]
- [=saltitans Scopoli, 1763: 350]
- [=spoliata Scopoli, 1763: 350]
- [=fasciata Schrank, 1803: 103]
- [=pallipes Say, 1823: 100]
- [=sulcicollis Meigen, 1824: 114]
- [=domesticus Meigen, 1824: 116]
- [=rufitarsis Meigen, 1838: 165]
- [=scutellatus Macquart, 1843: 6-8, plate 1]
- [=furcinervis Zetterstedt, 1844: 897]
- [="tarda" Linnaeus,- Haliday, 1851: 138]
- [=graminicola Zetterstedt, 1859: 6045]
- [=fuscinervis Schiner, 1862: 159]
- [=scutellatus var. nigroscutellatus Frey, 1945: 36]
- [=perkinsi Paramonov (not Hardy), 1955: 651]
- fijianus (Kröber) 1939: 399 (Omphrale), Australasian [velutinus group]
- flandersi Kelsey 1969: 132, Nearctic [velutinus group]
- flavipes (Kröber) 1939: 398 (Omphrale), Afrotropical [albicinctus group]
- foxi Kelsey 1971d: 9, Nearctic [velutinus group]
- fragosifrons Kelsey 1969: 68, Australasian [albicinctus group]
- fraternus (Kröber) 1923: 81 (Omphrale), Palearctic [brevicornis group]
- freidbergi Kelsey 1981: 10, Palearctic [albicinctus group]
- fryeri Kelsey 1969: 97, Afrotropical [brevicornis group]
- [=longiventris Lamb (not Krober) 1922: 362]
- [=minusculus Kelsey (not Seguy) 1969: 100]
- fulignatus Kelsey 1976: 953, Afrotropical [albicinctus group]
- galapagoensis Kelsey 1970: 96, Oceanian [velutinus group]
- gariesensis Kelsey 1976: 954, Afrotropical [albicinctus group]
- glabrifrons Meigen 1824: 114, Cosmopolitan
[fenestralis group]
- [=halterata Meigen 1824: 114]
- [=orbita Meigen 1824: 114]
- [=laevifrons Loew 1857: 88]
- [=glabrifrons var. surcoufi Séguy 1920: 318]
- [=civiculus G.H. Hardy 1933: 418]
- gobiensis Kelsey 1981: 89, Palearctic [brevicornis group]
- gossypius Kelsey 1969: 133, Nearctic [velutinus group]
- griseus (Kröber) 1913: 191 (Omphrale), Palearctic [fenestralis group]
- gromieri Kelsey 1969: 70, Afrotropical [albicinctus group]
- hagai Kelsey 1969: 135, Nearctic [velutinus group]
- halteralis Frey 1936: 53, Palearctic [brevicornis group]
- harleyi Kelsey 1971c: 187, Australasian [albicinctus group]
- hermonensis Kelsey 1981: 19, Palearctic [brevicornis group]
- herzliyanus Kelsey 1981: 12, Palearctic [albicinctus group]
- howdeni Kelsey 1969: 137, Nearctic [velutinus group]
- huldanus Kelsey 1981: 13, Palearctic [albicinctus group]
- ikoyianus Kelsey 1984: 54, Afrotropical [albicinctus group]
- inquilinus Séguy 1931: 113, Afrotropical [albicinctus group]
- japonicus (Séguy) 1920: 318 (Omphrale), Oriental [fenestralis group]
- johnsoni Kelsey 1971d: 9, Nearctic [velutinus group]
- josius Kelsey 1984: 55, Afrotropical [fenestralis group]
- kaszabi Kelsey 1981: 92, Palearctic [brevicornis group]
- katbergi Kelsey 1969: 72, Afrotropical [albicinctus group]
- keiseri Kelsey 1969: 74, Afrotropical [albicinctus group]
- kelseyi Ghorpade 1981: 53, Oriental [albicinctus group]
- kontagoranus Kelsey 1984: 56, Afrotropical [brevicornis group]
- kuiterti (D.E. Hardy) 1944a: 46, (Omphrale) Nearctic [velutinus group]
- lesinensis Strobl 1904: 533, Palearctic [fenestralis group]
- limbunyai Kelsey 1971c: 190, Australasian [albicinctus group]
- limpidipennis Loew 1874: 416, Palearctic [albicinctus group]
- lincinus Kelsey 1969: 99, Afrotropical [brevicornis group]
- linsleyanus Kelsey 1974b: 132, Nearctic [fenestralis group]
- longiventris (Kröber) 1913: 206 (Omphrale), Oriental [brevicornis group]
- lucidus Becker 1902: 39, Palearctic [albicinctus group]
- lunulanotafrons Kelsey 1971c: 191, Australasian [albicinctus group]
- maai Kelsey 1970a: 41, Oriental [albicinctus group]
- maculosus Kelsey 1981: 93, Palearctic [brevicornis group]
- madagascariensis Enderlein 1934: 429, Afrotropical [albicinctus group]
- magdalenai Kelsey 1969: 140, Nearctic [velutinus group]
- magnicornis (Kröber) 1913: 202 (Omphrale), Neotropical [velutinus group]
- majalcai Kelsey 1974b: 133, Neotropical [velutinus group]
- mariensis Kelsey 1981b: 88, Palearctic [fenestralis group]
- masoni Kelsey 1974b: 134, Nearctic [velutinus group]
- mendumae Kelsey 1971c: 188, Australasian [albicinctus group]
- microgaster (Séguy) 1948: 155 (Omphrale), Oriental [fenestralis group]
- millarae Kelsey 1971c: 189, Australasian [albicinctus group]
- minusculus (Séguy) 1934: 74 (Omphrale), Afrotropical [brevicornis group]
- minutus Kelsey 1969: 78, Australasian [albicinctus group]
- miriabilis Adams 1904: 445, Nearctic [velutinus group]
- [=nubilipes Cresson [nec Say] 1907: 111]
- [=ramaleyi James 1938: 22]
- monodi (Séguy) 1933: 122 (Omphrale), Afrotropical [albicinctus group]
- montanus Kelsey 1981: 14, Palearctic [albicinctus group]
- monterreyi Kelsey 1974b: 134, Nearctic [velutinus group]
- nakkari Kelsey 1987: 8, Australasian [albicinctus group]
- namibensis Kelsey 1976: 954, Afrotropical [fenestralis group]
- nidorcaupulus Kelsey 1969: 33, Nearctic [fenestralis group]
- niger (De Geer) 1776: 188 (Nemotelus), Palearctic [fenestralis
group]
- [=atra Fallén 1817: 2]
- [=fenestralis (Fabricius) 1781: 444]
- [=rugosa Fabricius 1794: 330]
- [=tarsata Panzer 1809: 20]
- [=nigripes Meigen 1824: 117]
- [=fasciata Walker 1851: 85]
- nigerianus Kelsey 1973a: 154 Afrotropical [fenestralis group]
- nitidifrons (Kröber) 1913: 194 (Omphrale) Afrotropical
[brevicornis group]
- [=tarsalis Kröber 1913: 196]
- nitidulus Loew 1873: 149, Palearctic [brevicornis group]
- niveus Becker 1907: 61, Palearctic [albicinctus group]
- norrisi Kelsey 1969: 82, Australasian [albicinctus group]
- nubilipes Say 1829: 170, Nearctic [velutinus group]
- obregoni Kelsey 1974b: 135, Nearctic [velutinus group]
- oldenbergi (Kröber) 1913: 190 (Omphrale), Palearctic [albicinctus group]
- opaculus Loew 1873: 145, Palearctic [fenestralis group]
- opacus De Meijere 1924: 14, Oriental [albicinctus group]
- orarius Kelsey 1981: 15, Palearctic [albicinctus group]
- pakhuisensis Kelsey 1976: 956, Afrotropical [albicinctus group]
- pallidipennis (Paramonov) 1955: 651 (Scenopinula), Australasian
- palmulapalpus Kelsey 1969: 85, Afrotropical [albicinctus group]
- papuanus (Kröber) 1912: 25 (Omphrale), Oriental, Australasian
[albicinctus group]
- [=niger Grimshaw (nec. De Geer) 1901: 11]
- [=zeylanicus Senior-White 1922: 205]
- parallelus Kelsey 1969: 103, Afrotropical [brevicornis group]
- patrizi (Séguy) 1932: 499 (Omphrale), Palearctic [albicinctus group]
- pecki Kelsey 1969: 145, Nearctic [velutinus group]
- peniculus Kelsey 1981: 16, Palearctic [albicinctus group]
- perkinsi G.H. Hardy 1942: 202, Australasian [albicinctus group]
- phaidimos Kelsey 1969: 88, Palearctic [albicinctus group]
- physadius (Séguy) 1930: 111 (Omphrale), Palearctic [fenestralis group]
- pilosus (Séguy) 1930: 111 (Omphrale), Palearctic [brevicornis group]
- planifrons Kelsey 1981: 20, Palearctic [brevicornis group]
- politus Kelsey 1973a: 156, Afrotropical [albicinctus group]
- pygmaeus Loew 1857: 90, Neotropical [velutinus group]
- [=albidipennis Loew 1869: 32]
- reciprocatus Kelsey 1971d: 10, Nearctic [velutinus group]
- reduncus Kelsey 1969: 106, Afrotropical [brevicornis group]
- renneri Kelsey 1975a: 204, Australasian [albicinctus group]
- retuertensis Carles-Tolrá 2001: 37, Palearctic [brevicornis group]
- rossi Kelsey 1969: 148, Nearctic [velutinus group]
- saini Kelsey 1969: 39, Nearctic [fenestralis group]
- sanfordi Kelsey 1971c: 192, Australasian [albicinctus group]
- schroederi (Kröber) 1913: 201 (Omphrale), Neotropical [fenestralis group]
- schulzi Enderlein 1934: 429, Neotropical [velutinus group]
- [=caenofrons Kröber 1937: 229]
- scintellatus Kelsey 1981: 22, Palearctic [brevicornis group]
- seftoni Kelsey 1969: 152, Nearctic [velutinus group]
- serratus Kelsey 1969: 153, Nearctic [velutinus group]
- sibiricus Krivosheina 1981: 162, Palearctic [fenestralis group]
- sigaloesis Kelsey 1981: 17, Palearctic [albicinctus group]
- sinensis (Kröber) 1928: 1 (Omphrale), Oriental [fenestralis group]
- spurrelli Kelsey 1969: 90, Australasian [albicinctus group]
- squamosus (Villeneuve) 1913: 111 (Omphralosoma), Palearctic [albicinctus group]
- stegmaieri Kelsey 1974b: 135, Neotropical [velutinus group]
- stuckenbergi Kelsey 1971a: 167, Afrotropical [brevicornis group]
- telleri Kelsey 1973a: 157, Afrotropical [albicinctus group]
- tetravirgulatus Kelsey 1981: 9, Palearctic [fenestralis group]
- thailandicus Kelsey 1970a: 41, Oriental [albicinctus group]
- torrensianus Kelsey 1980a: 7, Australasian [albicinctus group]
- transversus Kelsey 1971d: 3, Nearctic [fenestralis group]
- trigelasinus Kelsey 1971c: 194, Australasian [albicinctus group]
- triquetus Kelsey 1971c: 192, Australasian [albicinctus group]
- turneri Kelsey 1969: 107, Afrotropical [brevicornis group]
- umkomaasensis Kelsey 1976: 957, Afrotropical [albicinctus group]
- undulafrons Kelsey 1969: 41, Nearctic [fenestralis group]
- unifasciatus (Kröber) 1913: 195 (Omphrale), Palearctic [brevicornis group]
- ussuriensis Krivosheina 1981: 29, Palearctic [fenestralis group]
- valgus (D.E. Hardy) 1944a: 50 (Omphrale), Nearctic [velutinus group]
- vanduzeei Kelsey 1969: 156, Nearctic
- [=nubilipes Cole (nec. Say) 1923: 462]
- varipes Loew 1873: 148, Palearctic [fenestralis group]
- velutinus (Kröber) 1913: 203 (Omphrale), Neotropical
[velutinus group]
- [=nubilipes Kröber (nec. Say) 1928: 2]
- verrucosus Carles-Tolrá 2001: 36, Palearctic [brevicornis group]
- vesperugocavus Kelsey 1976: 958, Afrotropical [albicinctus group]
- vitripennis Meigen 1824: 115, Palearctic [fenestralis group]
- vockerothi Kelsey 1971d: 4, Nearctic [fenestralis group]
- wahrmani Kelsey 1981: 18, Palearctic [albicinctus group]
- weemsi Kelsey 1969: 159, Nearctic [velutinus group]
- werneri Kelsey 1969: 160, Nearctic [velutinus group]
- whittakeri (James) 1955: 47 (Omphrale), Nearctic [velutinus group]
- zambianus Kelsey 1973a: 158, Afrotropical [albicinctus group]
- zhelochovtsevi Krivosheina 1981: 160, Palearctic [fenestralis group]
- chinchona (Kelsey) 1969: 294 (Seguyella), Oriental
- deemingi Kelsey 1984: 59, Afrotropical
- galactica (Séguy) 1938: 334 (Omphrale), Afrotropical
- maai (Kelsey) 1970a: 46 (Seguyella), Oriental
- sinaiensis (Kelsey) 1981c: 24 (Seguyella), Palearctic
- turneri (Kelsey) 1969: 296 (Seguyella), Afrotropical
- flavoscutellata (Kröber) 1929: 80 (Pseudomphrale), Palearctic
- teutankhameni (Kröber) 1923: 72 (Pseudomphrale), Palearctic
- kirkspriggsi Winterton & Metz 2005 : 4, Afrotropical
- Subfamily Incertae Sedis
- kirkspriggsi Winterton & Metz 2005 : 4, Afrotropical
- Dichotomous key to subfamilies genera
- Scenopinid Literature
- Adams, C.F. (1904) Notes on and descriptions of North American Diptera. Kansas University Science Bulletin 2: 433-455.
- Agassiz, L. (1846) Nomenclatoris Zoologici. Index Universalis. pp. 393 Solodure, Switzerland.
- Alexander, K.N.A. (1991) Scenopinus niger DeGeer (Scenopinidae) in Montgomeryshire. Dipterists Digest 8: 24.
- Alexander, K.N.A. (1996) A review of records for Scenopinus niger (Degeer) (Diptera, Scenopinidae) in Wales. Dipterists Digest Second Series 2: 90-91.
- Allen, A.A. (1979) An outdoor occurrence of Scenopinus fenestralis (L.) (Dipt., Scenopinidae). Entomologists Monthly Magazine 115: 200. [text states that a pair of specimens swept from foliage in garden]
- Arnaud, P.H. (1979) A catalog of the types of Diptera in the collection of the California Academy of Sciences. Myia 1: 139-150.
- Becker, T. (1902) Aegyptische Dipteren. Berlin Zool. Museum Mitt. 2: 1-66, [+1 plate].
- Becker, T. (1907) Die Ergebnisse meiner dipterologischen Frühjahrsreise nach Algier und Tunis, 1906 (Concl.) Z. system. Hym. Diptera 7: 33-61, 97-128, 225-256, 369-407, 454, [+12 figures].
- Becker, T. (1908) Dipteren der Kanarischen Inseln. Mitteilungen aus dem zoologischen Museum in Berlin 4(1): 1-180; Berlin.
- Becker, T. (1913) Persische Dipteren von den Expeditionen des Herrn N. Zarudny, 1898 und 1901. Annals Museum of Zoology Academy of Science St. Petersburg 17: 503-654. (1912) [+3 plates].
- Bezzi, M. (1922) On the South American species of the Dipterous genus Chriomyza Wied. (Diptera). Annals of the Entomological Society of America 15: 117-124.
- Bezzi, M. (1925a) Une Novelle Espece du Genre Pseudomphrale de l'Afrique du Sud. Encyclopedie Entomologique. Series B. Diptera 2: 95-98.
- Bezzi, M. (1925b) Quelques notes sur les Bombyliides (dipt.) d'Egypte, avec desciption d'espèces nouvelles. Bulletin de la Société Entomologique d'Egypte 8:159-242. [1924]
- Bosak, J. (1996) Faunistic records from the Czech Republic - 52. Diptera: Scenopinidae. Klapalekiana 32: 270.
- Bosak, J. (1997) Scenopinidae (Diptera) of the Czech and Slovak Republics. Folia Facultatis Scientiarum Naturalium Universitatis Masarykianae Brunensis Biologia 95: 23-25.
- Bosak, J. & Bartak, M. (2000) Scenopinidae. Folia Facultatis Scientiarum Naturalium Universitatis Masarykianae Brunensis Biologia 104: 163-165.
- Bowden, J. (1999) Scenopinidae (Dipt.) in N.E. Essex. Entomologists Monthly Magazine 135: 176. [Records Sc. fenestralis & Sc. niger in Essex]
- Brauns, A. (1954) Untersuchungen zur angewandten Bodenbiologie. Vol. 2: Puppen terricole Dipterenlarven. Gottingen, Frankfurt, Berlin. 156 pp.
- Carles-Tolrá, M. (1995) Insecta: Diptera, 20-26. Families 20-26: Conopidae, Xenasteiidae, Acroceridae, Nemestrinidae, Scenopinidae, Stratiomyidae and Vermileonidae. Catalogus de la Entomofauna Aragonesa 8: 7-9
- Carles-Tolrá, M. (1998) New data on the Diptera from Spain (Diptera: Scenopinidae, Xylomyidae, Lonchopteridae, Hippoboscidae, Nycteribiidae). Boletin de la Asociacion Espanola de Entomologia 22: 75-79.
- Carles-Tolrá, M. (1999a) New and interesting Diptera to the Iberian Peninsula and Balearic Islands (Diptera: Xylomyidae, Stratiomyidae, Acroceridae, Scenopinidae, Lonchopteridae, Opetiidae, Platypezidae, Conopidae). Boletin de la Asociacion Espanola de Entomologia 23: 195-212.
- Carles-Tolrá, M. (1999b) Scenopinus efflatouni Kelsey, 1969: new record to Europe (Diptera, Scenopinidae). Boletin de la Asociacion Espanola de Entomologia 23: 328.
- Carles-Tolrá, M. (2001) Two new species of Scenopinus Latreille from Spain (Diptera, Scenopinidae). Boletin de la Asociacion Espanola de Entomologia 25: 35-41.
- Cole, F.R. (1923a) A revision of the North American two-winged flies of the family Therevidae. Proceedings of the United States National Museum 62: 1-140 [+13 plates].
- Cole, F.R. (1923b) Expedition of the California Academy of Sciences to the Gulf of California in 1921. Diptera from the islands and adjacent shores of the Gulf of California. II. General report. Proceedings of the California Academy of Sciences 12: 457-481.
- Cole, F.R. & Schlinger, E.I. (1969) Scenopinidae. In F.R. Cole The flies of western North America. University of California Press, Berkeley.
- Coquillett, D.W. (1900) New Scenopinidae from the United States. Entomological News 11: 500-501.
- Coquillett, D.W. (1902) New Diptera from North America. Proceedings of the U.S. National Museum 25: 83-126.
- Cresson, E.T. (1907) The North American species of the dipterous family Scenopinidae. Transactions of the American Entomological Society 33: 109-114.
- Curran, C.H. (1928) Diptera of the American Museum Congo Expedition. Part 3, Stratiomyidae, Rhagionidae, Therevidae, Scenopinidae, Ortalidae, Micropezidae, Piophilidae, Sepsidae, and Diopsidae. Bulletin of the American Museum of Natural History 58 (4): 167-187.
- Dalla Torre, K.W. (1877 [1878]) Beitrage zur Phyto-& Zoostatik des Egerlandes. Jehresb. naturh. Ver. Lotos 27: 7-208.
- De Geer, C. (1776) Mémoires pour servir à l'histoire des Insectes. 5: 1-523. [30 plates] Stockholm.
- De Meijere, J.C.H. (1924) Studien über Südostasiatische Dipteren, XV. Tijdschrift voor Entomologie 67: 1-64.
- Daniels, G. (1978) Type specimens of Diptera in the Australian Museum. Records of the Australian Museum 31: 411-471.
- Dobson, J.R. (1999) Does Scenopinus fenestralis (Linnaeus) (Diptera, Scenopinidae) breed in beehives? Dipterists Digest Second Series 6: 42.
- Dunk, K. (1995) Zweifluger aus Bayern 6 (Diptera: Acroceridae, Scenopinidae, Pipunculidae). Entomofauna 16: 373-380.
- Edwards, F.W. (1932) Scenopinidae, Mydaidae. Diptera of Patagonia and south Chile. Part 5, fasc. 3, pp. 258-260.
- Efflatoun, H.C. (1945) A monograph of Egyptian Diptera. Part IV. Family Bombyliidae. Section I: Subfamily Bombyliidae Homeophthalmae. Bulletin de la Societe Fouad Ier d'Entomologie 29: 1-483.
- Enderlein, G. (1914) Dipterologische Studien X. Zur Kenntnis der Stratiomyiiden mit 3 zästiger Media und ihre Gruppierung. B, Formen, bei denen der 1. Cubitalast mit der Discoidalzelle eine Strecke verschmolzen ist (Familien: Hermetiinae, Clitellariinae). Zoologisher Anzeiger 44: 1-25.
- Enderlein, G. (1934) Dipterologica. I. Sber. Ges. Naturf. Freunde Berl. 1933: 416-429.
- Engel, E.O. (1932) Family Omphralidae. pp:123-126. In F. Dahl, (ed.) Die Tierweldt Deutschlands vol. 26, teil 5, pp. 204, Jena.
- Evenhuis, N.L. (1980) Catalog of entomological types in the Bishop Museum. Diptera: Rachiceridae, Scenopinidae, Empididiae. Pacific Insects 22: 212-215.
- Evenhuis, N.L. (1991) World catalog of genus-group names of bee flies (Diptera: Bombyliidae). Bishop Museum Bulletin Entomol. 5: 1-105.
- Fabricius, J.C. (1794) Entomologia systematica emendata et aucta. Secundum classes, ordines, genera, species adjectis synonimis, locis, observationibus, descriptionibus. Vol 4, [6]+ 472+ [5] pp. C.G. Proft, Hafniae [=Copenhagen].
- Fallén, C.F. (1817) Observationes Entomologicae. Scenopinii et Conopsariae Sveciae. pp. 14. Lundae.
- Frey, R. (1936) Die Dipteren fauna der Kanarischen Inseln und ihre Probleme. Societas Scientiarum Fennica. Commentiones Biologicae 6: 1-237. [+10 plates]
- Frey, R. (1945) Tiergeographische Studien uber die Dipteren fauna der Azoren. 1. Verzeichnis der bisher von den Azoren bekannten Dipteren. Commentat. Biol. Soc. Sci. Fenn. (1944) 8(10): 114 p.
- Ghorpade, K.D. (1981) On a new species of Scenopinus (Diptera: Scenopinidae) from south India. Colemania 1: 53-54.
- Gnaspini, N,P. (1989) Comparative analysis of fauna associated with guano deposits of cavernicolous bats in Brazil, First Estimate. Revista Brasileira de Entomologia 33: 183-192. [Sc. fenestralis recorded from guano]
- Grimaldi, D. & Cumming, J. (1999) Brachyceran diptera in cretaceous ambers and mesozoic diversification of the Eremoneura. Bulletin of the American Museum of Natural History 239: 1-124. [8 plates]
- Haliday, A.H. (1851) Über die Dipteren der in London befindlichen Linnéischen Sammlung. Stettiner Entomologische Zeitung 12: 131-145.
- Hall, J.C. (1972) New North American Heterotropinae (Diptera: Bombyliidae). The Pan-Pacific Entomologist 48: 37-50.
- Hardy, D.E. (1944a) A revision of the North American Omphralidae (Scenopinidae). Journal of the Kansas Entomological Society 17: 31-40, 42-51.
- Hardy, D.E. (1944b) A new Pseudatrichia from Brazil (Omphralidae- Scenopinidae). Kansas Entomological Society Journal 17: 104-105.
- Hardy, D.E. (1958) Insects of Micronesia Diptera: Omphralidae (Scenopinidae), vol. 13: 11-13.
- Hardy, D.E. (1960) Diptera: Nematocera-Brachycera (except Dolichopodidae). In E.C. Zimmerman (ed.) Insects of Hawaii, Honolulu. vol 10. pp. 368.
- Hardy, D.E. (1966) Family Scenopinidae (Omphralidae). In N. Papavero, (ed.) A Catalogue of the Diptera of the Americas south of the United States, pp: 32.1-32.5, Departamento de Zoologia, Secretaria da Agricultura. São Paulo.
- Hardy, D.E. (1983) Family Scenopinidae (Omphralidae) In A. Stone, C.W. Sabrowsky, W.W. Wirth, R.M. Foote & J.R. Caulson (eds), A Catalogue of Diptera of North America, pp 354-356, Smithsonian Publication, Washington D.C.
- Hardy, G.H. (1933) Miscellaneous notes on Australian Diptera, I. Proceedings of the Linnaean Society of New South Wales 58: 408-420.
- Hardy, G.H. (1942) Miscellaneous notes on Australian Diptera, IX: Superfamily Asiloidea. Proceedings of the Linnaean Society of New South Wales 67: 197-204.
- Hassan, S.A. & El Hawagry, M.S.A (2001) A revision of the family Scenopinidae (Diptera) from Egypt. Efflatounia 1: 1-11.
- Hendel, F. 1908. Nouvelle classification des mouches à deux ailes (Diptera L.). D'après un plan tout nouveau par J.G. Meigen, Paris, an VIII (1800 vés.). Verh. Zool.-Bot. Ges. Wien 58: 43-69.
- Hull, F.M. (1973) Bee flies of the world. The genera of the family Bombyliidae. Bulletin of the United States National Museum 286: 1-687.
- International Commission on Zoological Nomenclature (1963) Opinion 678. The suppression under the plenary powers of the pamphlet published by Meigen, 1800. Bulletin of Zoological Nomenclature 20: 339.
- James, M.T. (1938) The dipterous families Nemestrinidae, Cyrtidae and Scenopinidae in Colorado. Journal of the Kansas Entomological Society 11: 21-23.
- James, M.T. (1955) Two new Diptera from the Pacific coast states. Journal of the Kansas Entomological Society 28: 47-48.
- Kelsey, L.P. (1969) A revision of the Scenopinidae (Diptera) of the world. Bulletin of the United States National Museum 277: 1-336.
- Kelsey, L.P. (1970a) New Scenopinidae (Diptera) from the Pacific area. Pacific Insects 12: 39-48.
- Kelsey, L.P. (1970b) The Scenopinidae (Diptera) of Australia: including the description of one new genus and six new species. Journal of the Australian Entomological Society 9: 103-148.
- Kelsey, L.P. (1970c) A new Scenopinus from the 1964 Galapagos International Scientific Project (Diptera Scenopinidae). The Pan-Pacific Entomologist 46: 96-97.
- Kelsey, L.P. (1971a) A review of the Scenopinidae (Diptera) of Africa south of the equator and neighbouring islands, with descriptions of two new species. Annals of the Natal Museum 21: 135-181.
- Kelsey, L.P. (1971b) A new scenopinid genus with three new species from Chile. The Pan-Pacific Entomologist 47: 279-284.
- Kelsey, L.P. (1971c) Eighteen new species of Australian Scenopinidae (Diptera). Journal of the Australian Entomological Society 10: 183-204.
- Kelsey, L.P. (1971d) New Scenopinidae (Diptera) from North America. California Academy of Sciences, Occasional Papers 88: 1-65.
- Kelsey, L.P. (1971e) A new Scenopinidae (Diptera) from Bermuda. Psyche 78: 49-50.
- Kelsey, L.P. (1973a) New Scenopinidae (Diptera) from the Ethiopian region. Journal of Natural History 7: 153-160.
- Kelsey, L.P. (1973b) The zoogeographic distribution of known Scenopinidae (Diptera). Entomological News 84: 329-332.
- Kelsey, L.P. (1974a) A description of the male of Scenopinus politus Kelsey. Journal of Natural History 8: 153-154.
- Kelsey, L.P. (1974b) Eleven new Scenopinidae (Diptera) from the Americas. Quaestiones Entomologicae 10: 131-147.
- Kelsey, L.P. (1975a) New Australian Scenopinidae (Diptera). Journal of the Australian Entomological Society 14: 197-212.
- Kelsey, L.P. (1975b) Family Scenopinidae. In M.D. Delfinado & D.E. Hardy (eds), A Catalog of the Diptera of the Oriental Region. vol. II. Suborder Brachycera- Division Aschiza, Suborder Cyclorrhapha. pp. 94-95. University Press Hawaii, Honolulu.
- Kelsey, L.P. (1976) New African Scenopinidae (Diptera) from south of the equator. Annals of the Natal Museum 22: 951-977.
- Kelsey, L.P. (1980a) New Australian Scenopinidae (Diptera) III. Journal of the Australian Entomological Society 19: 7-10.
- Kelsey, L.P. (1980b) Family Scenopinidae. In R.W. Crosskey (ed.), Catalogue of the Diptera from the Afrotropical region, pp 321-323. British Museum (Natural History), London.
- Kelsey, L.P. (1981a) Scenopinidae. In J.F. McAlpine, B.V. Peterson, G.E. Shewell, H.J. Teskey, J.R. Vockeroth & D.M. Wood (eds), Manual of Nearctic Diptera 1: 525-528. Research Branch, Agriculture Canada Monograph, Ottawa 674 pp.
- Kelsey, L.P. (1981b) New Scenopinidae (Diptera) from the Palearctic. Folia Entomologica Hungarica 2: 85-93.
- Kelsey, L.P. (1981c) Scenopinidae (Diptera) of Palestine and the Sinai peninsula. Entomologists Monthly Magazine 117: 3-25.
- Kelsey, L.P. (1984) New Scenopinidae (Diptera) from Nigeria. Nigerian Journal of Entomology 5: 50-61.
- Kelsey, L.P. (1987) New Australian Scenopinidae (Diptera) IV. Journal of the Australian Entomological Society 26: 1-32.
- Kelsey, L.P. (1989a) Family Scenopinidae. In N.L. Evenhuis (ed.) Catalog of Diptera of the Australasian and Oceanian Regions. pp. 350-352. Bishop Museum Special Publication. Bishop Museum Press 86: 1-1154.
- Kelsey, L.P. & Soos, A. (1989b) Family Scenopinidae. In A. Soos & L. Papp (eds.) Catalogue of Palearctic Diptera . Therevidae-Empididae, 6: 35-43. Akadémial Kiadó, Budapest.
- Kertész, K. (1899) Verzeichniss einiger, von L. Biró in Neu-Guinea und am Malayischen Archipel. Gesmmelten Dipteren. Természetrajzi Fuzetek 22: 173-195.
- Krivosheina, N.P. (1980) New scenopinids (Diptera: Scenopinidae) from the Palearctic. Entomologicheskoe Obozrenie 59: 197-205.
- Krivosheina, N.P. (1981a) Taxonomy of species of the Scenopinus fenestralis group (Diptera: Scenopinidae). Vestnik Zoologii 4: 24-31.
- Krivosheina, N.P. (1981b) New representatives of the genus Scenopinus (Diptera: Scenopinidae) in the fauna of the USSR. Zoologicheskii Zhurnal 60: 160-164.
- Krivosheina, N.P. & Krivosheina, M.G. (1996a) Description of the type specimens of dipterans of the genus Pseudomphrale Kröber (Diptera, Scenopinidae). Entomological Obozrenie 76: 494-498.
- Krivosheina, N.P. & Krivosheina, M.G. (1999) New data on Palearctic species of the genus Metatrichia (Diptera: Scenopinidae). Zoologicheskii Zhurnal 78: 849-859.
- Krivosheina, N.P. (1997) Family Scenopinidae. In L. Papp & B. Darvas (eds) Contributions to a manual of Palearctic Diptera (with special reference to flies of economic importance). Volume 2: Nematocera and lower Brachycera. pp. 531-538. Science Herald, Budapest.
- Kröber, O. (1912) H. Sauter's Formosa-Ausbeute. Thereviden, Omphraliden (Dipt.). Suppl. Ent. 1: 24-26.
- Kröber, O. (1913) Die Omphraliden. Eine Monographische Studie. Annales Museum naturale Hungary 11: 174-210.
- Kröber, O. (1914a) Diptera, Fam. Omphralidae. Fasc. 161. pp. 16 [+4 plates]. In P. Wytsman (ed.) Genera Insectorum Bruxelles.
- Kröber, O. (1914b) Beiträge zur Kenntnis der Thereviden und Omphraliden. Jahrbuch der Hamburgischen Wissenschaftlichen Anstalten (1913) 31: 29-74.
- Kröber, O. (1923) Aegyptische Dipteren aus den Familien der Conopidae, Omphralidae und Therevidae. Bulletin de la Société Royale Entomologique d'Egypte 7: 57-116.
- Kröber, O. (1925) Omphralidae (Scenopinidae). In E. Lindner (ed.) Die Fliegen der Palaearktischen Region. Vol. 4. pt 3 sect 2. fam 27. pp. 1-8. 2 plates.
- Kröber, O. (1928a) Neue and wenig bekannte Diptern aus den Familien Omphralidae, Conopidae und Therevidae. Konowia 7: 1-23.
- Kröber, O. (1928b) Neue Dipteren des Deutschen Entomolog. Museums in Dahlem (Conopidae, Omphralidae, Therevidae, Tabanidae). Entomologische Mitteilungen 17: 31-41.
- Kröber, O. (1929) Neue Dipteren aus Aegypten aus den Familien Tabanidae, Therevidae, Omphralidae u. Conopidae. Bulletin de la Société Entomologique d'Egypte 13: 73-84.
- Kröber, O. (1937) Ein Beitrag zur Kenntnis der Omphraliden (Scenopinidae), Diptera. Stettiner Entomologische Zeitung 98: 211-231.
- Kröber, O. (1939) Beitrag zur Kenntnis der Therevidae und Omphralidae. Annals and Magazine of Natural History 3: 395-400.
- Lamb, C.G. (1922) The Percy Sladen Trust Expedition to the Indian Ocean in 1905. No. VIII. Diptera: Asilidae, Scenopinidae, Dolichopodidae, Pipunculidae, Syrphidae. Transactions of the Linnaean Society of London 18: 361-416.
- Latreille, P.A. (1802) Histoire Naturelle, générale et particulière des Crustacés et des Insectes. Tome troisième. Families naturelles et genres. In C.S. Sonnini (ed.) Histoire Naturelle par Buffon, Paris, pp. 468.
- Lindner, E. (1955) Ostafrikanische, Omphralidae, Therevidae und Conopidae (Dipt.) Jahreshefte Vereins fur Vaterlaendische Naturkunde in Wuerttemburg 110: 19-23.
- Lindner, E. (1962) Afrikanische Rhagionidae, Therevidae und Omphralidae, (Dipt.) Stuttgarter Beitraege zur Naturkunde 83: 1-3.
- Linnaeus, C. (1758) Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum caracteribus, differentiis, synonymis, locis. 10th Edition, vol. 1, 824 pp. L. Salvii, Holmiae [=Stockholm].
- Linnaeus, C. (1761) Fauna Svecica sistens Animalia Sveciae Regni: Mammalia, Aves, Amphibia, Pisces, Insecta, Vermes. Distributa per classes & ordines, genera & species, cum differentiis specierum, synonymis auctorum, nominibus incolarum, locis natalium, descriptionibus insectorum. Editio altera, auctior [49]+ 578 pp. 2 plates. L. Salvii, Stockholmiae.
- Lioy, P. (1864) I ditteri distribuiti secundo un nuova metodo di classificazione naturale. Atti Ist. Neneto Sci., series 3, vol. 9: 499-518, 569-604, 719-771, 879-910, 989-1027, 1311-1352.
- Liu, N. & Nagatomi, A. (1995) The mouthpart structure of Scenopinidae (Diptera). Japanese Journal of Entomology 63: 181-202.
- Loew, H. (1845) Ueber die systemat. Stellung von Scenopinus, und Beschreibung einer neuen Art. Stettiner Entomologische Zeitung 6: 312-315. [Sc. zelleri description p. 314-315]
- Loew, H. (1857) Die bis jetzt bekanntgewordenen Arten der Gattung Scenopinus. Verhandl. Zool. -Bot. Ver. Wien, 7: 87-90.
- Loew, H. (1866) Diptera Americae septentrionalis indigena. Centuria septima. Berliner Entomologische Zeitschrift 10: 1-54.
- Loew, H. (1869) Diptera Americae septentrionalis indigena. Centuria octava. Berliner Entomologische Zeitschrift 13: 1-52.
- Loew, H. (1873) Beschreibung europäischer Dipteren. Systematische Beschreibung der bekannten europäischen zweiflügeligen Insecten, von Johann Wilhelm Meigen. vol. 3, Zehnter Theil oder vierter Supplementband, 320 pp. Halle.
- Loew, H. (1874) Diptera nova a Hug. Theod. Christopho Collecta. Ztschr. f. die Gesam. Naturw., 43: 413-420.
- Lyneborg, L. (1965) Scenopinidae of Danmark (keys). Danmark Fauna 70: 77-80.
- Macquart, J. (1835) Histoire naturelle des insectes. Dipteres. Tome deuxieme. Roret, Paris. 703 or 710 pp., 12 pls.
- Macquart, J. (1843) Diptères exotiqes nouveaux ou peu connus. Vol. 2, pp. 5-304 [+36 plates]. Paris.
- Marsden, M.A., Furniss, M.M. & Kline, L.N. (1981) Modeling seasonal abundance of Douglas-fir beetle in relation to entomophagous insects and location in trees. US Forest Service General Technical Report INT. 1981: 1-22; No. 111.
- Meigen, J.G. (1800) Nouvelles classification des mouches à deux ailes (Diptera L.) d'après un plan tout nouveau. pp.40 , Paris. [suppressed, see ICZN (1968)]
- Meigen, J.G. (1803) Versuch einer neuen Gattungseintheilung der europäischen zweiflügeligen Insekten. Mag. Insektenkunde 2: 259-281.
- Meigen, J.G. (1824) Systematische Beschreibung der bekannten europäischen zweiflügeligen Insekten. Vierter Theil. Schulz-Wundermann, Hamm.xii + 428 pp. [+9 plates]
- Meigen, J.G. (1838) Systematische Beschreibung der bekannten europaischen zweiflugeligen Insekten. Siebenter Theil oder Supplementband. Schultz, Hamm. xii + 434 + [1] pp.
- Melander, A.L. (1906) Some new or little-known genera of Empididae. Entomological News 17: 370-379.
- Melander, A.L. (1950) Taxonomic notes on some smaller Bombyliidae (Diptera). The Pan-Pacific Entomologist 26: 139-156.
- Metz, M.A., Winterton, S.L. & Irwin, M.E. (2002) Notes on the functional morphology of terminalia from Prorates ballmeri Nagatomi & Liu (Diptera: Scenopinidae: Proratinae) collected while in copula, with a description of the previously unknown female. Zootaxa 76: 1-12.
- Metz, M.A. (2003) Description of a new species of Caenotus Cole (Diptera: Scenopinidae) from Baja California Sur, Mexico, with a review of the genus. Zootaxa 223: 1-11.
- Nagatomi, A., Saigusa, T., Nagatomi, H. and Lyneborg, L. (1991a) Apsilocephalidae, a new family of the orthorrhaphous Brachycera (Insecta, Diptera). Zoological Science 8: 579-591.
- Nagatomi, A., Saigusa, T., Nagatomi, H. & Lyneborg, L. (1991b) The systematic position of the Apsilocephalidae, Rhagionempididae, Protempididae, Hilarimorphidae, Vermileonidae and some genera of Bombyliidae (Insecta, Diptera) Zoological Science 8: 593-607.
- Nagatomi, A., Saigusa, T., Nagatomi, H. & Lyneborg, L. (1991c) The genitalia of Apsilocephalidae. Japanese Journal of Entomology 59: 409-423.
- Nagatomi, A., Liu, N. & Yanagida, K. (1994) Notes on the Proratinae (Diptera: Scenopinidae). South Pacific Study 14: 137-222.
- Nagatomi, A., Liu, N., & Evenhuis, N.L. (1994) The genitalia of Scenopinus glabrifrons (Diptera: Scenopinidae). Japanese Journal of Entomology 62: 1-12.
- Narchuk, E.P. (1988) Family Scenopinidae (Omphralidae). In Bei Bienko, G.Ya. (ed.) Keys to the insects of the European part of the USSR. Volume 5: Diptera and Siphonaptera. Part 1. pp: 838-842. Smithsonian Institution Libraries & National Research Foundation. Washington, DC. i-xxii, 1-1233.
- Osten Sacken, C.R. (1877) Western Diptera: Descriptions of new genera and species of Diptera from the region west of the Mississippi and especially from California. Bulletin of the United States Geological and Geographical Survey of the Territories 3: 189-354.
- Ovtshinnikova, O.G. & Yeates, D.K. (1998) Male genital musculature of Therevidae and Scenopinidae (Diptera: Asiloidea): Structure, homology and phylogenetic implications. Australian Journal of Entomology 37: 27-33.
- Panzer, G.W.F. (1809) Favnae insectorvm germanicae initia oder Devtschlands Insecten. H. 108. Felsecker, Nurnberg [= Nuremberg]. 24 pp. [+24 plates]
- Paramonov, S.J. (1929) Beiträge zur Monographie einiger Bombyliiden-Gattungen (Diptera). Zb. Pra. Zool. Muz. 6: 1-161.
- Paramonov, S.J. (1955) A review of Australian Scenopinidae (Diptera). Australian Journal of Zoology 1: 634-653.
- Qvick, U. (1983) Aerial swarming in Scenopinus niger Degeer (Scenopinidae). Notulae Entomologicae 63: 216.
- Rafael, J.A. & Ale, R.A. (1983) Ocorréncia de Metatrichia robusta Kröber (Diptera: Scenopinidae) na Amazônia e sua Distribução sazonal. Acta Amazonica 13: 709-711.
- Rahman, H.A.A., Shaumar, N.F., Soliman, Z.A. & El Agoze, M.M. (1977 [1981]) Survey and taxonomy of parasites and predators of stored grain and grain products insects. Bulletin de la Société Entomologique d'Egypte 61: 53-74.
- Rondani (1848) Esame di varie specie d'insetti ditteri brasiliani. Stud. Ent. (Turin) 1: 63-112.
- Rossi, P. (1794) Mantissa Insectorum, exhibens species nuper in Etruria collectas, adjectis faunae Etruscae illustrationibus a c emendationibus. Pisa. Polloni, 1792-1794, 4. 2 vol. 148 pp & 154 pp.
- Say, T. (1823) Descriptions of dipterous insects of the United States. Journal of the Academy of Natural Sciences Philadelphia 3: 9-54, 73-104.
- Say, T. (1829) Descriptions of North American dipterous insects. Journal of the Academy of Natural Sciences Philadelphia 6: 149-178.
- Schellenberg, J.R. (1803) Genres des mouches Diptères représentés en XLII planches projettées et dessinées et expliquées par deux amatuers de l'entomologie. Zurich. 95 pp., 42 plates.
- Schembri, S., Gatt, P. & Schembri, J. (1991) Recent records of flies from the Maltese Islands: Diptera. Memorie della Societa Entomologica Italiana 70: 255-278.
- Schiner, I.R. (1862) Die Fliegen (Diptera). In L. Redtenbacher & I.R. Schiner (eds) Fauna Austriaca vol. 1, 674 pp. [2 plates]
- Schrank, F. P. (1803) Favna Boica. Durchgedachte Geschichte der in Baiern einheimischen und zahmen Thiere. Vol. 3, Pt. 1. P. Krull, Landshut. viii + 272 pp.
- Scopoli, J.A. (1763) Entomologia carniolica carniolica exibens insecta carniolica indigena et distributa in ordines, genera, species, varietates, methodo Linnaeana. I. T. Trattner, Vindobonae [=Vienna]. [36] + 420 + [1] pp.
- Séguy, E. (1920) Notes sur quelques Omphrale (Dipt. Omphralidae) et descriptions d'especes nouvelles. Bulletin Society Entomologique de France 1920: 312-319.
- Séguy, E. (1921) Étude sur L'Omphrale fenestralis L. (Diptera). Bull. Mus. Hist. Nat. Paris 27: 60-66.
- Séguy, E. (1926) Diptères (Brachyceres). Fauna de France 13: 275-279.
- Séguy, E. (1930) Contribution à l'étude des dipteres du Maroc. Mem. Soc.Sci. Nat. Maroc 24, 206 pp.
- Séguy, E. (1931) Contribution a l'étude de la Faune du Mozambique. Voyage de M.P. Lesne 1928-1929. Bulletin du Museum d'Histoire Naturelle (Paris) 3: 113-121.
- Séguy, E. (1932) Spedizione scientifica all'oasi di Cufra (Marzo-Luglio 1931) Insectes Diptères. Ann. Mus. Stor. Nat. Genova 55: 490-511.
- Séguy, E. (1933) Mission Saharienne Augiéras-Draper 1927-1928. Insectes Diptères. Bulletin du Museum d'Histoire Naturelle (Paris) sér. 2, vol. 5: 122-127.
- Séguy, E. (1934) Diptera d'Afrique. Encyclopedie Entomologique. Series B. Diptera 7: 63-80.
- Séguy, E. (1938) Mission scientifique de l'Omo. Diptera 1, Nematocera et Brachycera. Memoirs of the Museum of Natural History, Paris 8: 319-380.
- Séguy, E. (1948) Dipteres nouveaux ou peu connus d'extreme-Orient. Notes Ent. Chin. 12(14): 153-172.
- Séguy, E. (1968) Remarks on some dipterous insects of the genus Omphrale (Diptera Scenopinidae), Omphrale provancheri new species, Omphrale minuscula female 1st description. Naturaliste Canadien Quebec. 95: 657-659.
- Senior-White, R.A. (1922 [1924]) New Ceylon Diptera. Spolia Zeylan. 13: 193-283, 12 plates.
- Speiser, P. (1920) Zur Kenntnis der Diptera Orthorrhapha Brachycera. Zoologische Jahrbucher 43: 195-220.
- Strobl, G. (1904) Neue Beiträge zur Dipterenfauna der Balkanhalbinsel. Wissenschaftliche Mittheilungen aus Bosnien und der Herzegowina 9: 519-581.
- Stubbs, A.E. & Drake, M. (2001) British soldierflies and their allies. An illustrated guide to their identification and ecology, covering all flies (Diptera) in the families Acroceridae, Asilidae, Athericidae, Bombyliidae, Rhagionidae, Scenopinidae, Stratiomyidae, Tabanidae, Therevidae, Xylomyidae and Xylophagidae. British Entomological and Natural History Society 2001: [x]+ 1-512.
- Theodor, O. (1983) The genitalia of Bombyliidae (Diptera). Jeruselum: The Israel Academy of Sciences and Humanities. pp. 275.
- Thompson, J.H., Knutson, L.V. & Culp, O.S. (1970) Larva of Scenopinus sp. (Diptera Scenopinidae) causing human uro-genital myiasis. Mayo Clinic Proceedings 45: 597-601.
- Trojan, P. (1956a) Notes on the taxonomy of some European species of the genus Omphrale Meigen (Diptera: Omphralidae). Annales Zoologici Pan. 16: 147-156.
- Trojan, P. (1956b) Omphralidae. Klucze do Oznaczania Owadow Polskich 28: 3-15.
- Van Duzee, M.C. (1926) A new species of Scenopinidae from California (Diptera). The Pan-Pacific Entomologist 2: 164.
- Villeneuve, J. (1913) Diptères nouveaux ou intéressants. Feuille des Jeunes Naturalistes 43: 111-113.
- Walker F. (1851) In F. Walker, H.T. Stainton & S.J. Wilkinson, (eds) Insecta Britannica Diptera vol. 1, 314 pp. [10 plates] London.
- Winterton S.L. (2005) A new species of Propebrevitrichia Kelsey (Diptera: Scenopinidae: Scenopininae) from Botswana. Zootaxa 818: 1-8.
- Winterton, S.L. & P.H. Kerr (2006) A new species of Alloxytropus Bezzi (Diptera: Scenopinidae: Proratinae) from Israel. Zootaxa 1155: 41-50.
- Winterton, S.L. & Metz, M.A. (2005) Cyrtosathe gen. n., the first non-scenopinine window-fly from sub-Saharan Africa (Diptera: Scenopinidae). Zootaxa 975: 1-12.
- Woodley, N. & Hilburn, D.J. (1994) The Diptera of Bermuda. Contributions of the American Entomological Institute 28: 1-64.
- Yang, L., Wiegmann, B.M., Yeates, D.K. & Irwin, M.E. (2000) Higher-level phylogeny of the Therevidae (Diptera: Insecta) based on 28S ribosomal and elongation factor-1alpha gene sequences. Molecular Phylogenetics and Evolution 15: 440-451.
- Yao, M.C. & Lo, K.C. (1992) Insect species and population densities in stored japonica rice in Taiwan. Chinese Journal of Entomology 12: 161-169. [Sc. fenestralis records in stored rice]
- Yeates, D.K. (1992) Towards a monophyletic Bombyliidae (Diptera): the removal of the Proratinae (Diptera: Scenopinidae). American Museum Novitates 3051: 1-30.
- Yeates, D.K. & Grimaldi, D. (1993) A new Metatrichia Window Fly (Diptera: Scenopinidae) in Dominican Amber, with a review of the systematics and biogeography of the genus. American Museum Novitates 3078: 1-8.
- Yeates, D.K. (1994) The cladistics and classification of the bombyliidae (Diptera: Asiloidea). Bulletin of the American Museum of Natural History. 219: 1-191.
- Yeates, D.K. (2002) Relationships of extant lower Brachycera (Diptera): A quantitative synthesis of morphological characters. Zoologica Scripta 31: 105-121.
- Yeates, D.K, Irwin, M.E. & Wiegmann, B.M. (2003) Ocoidae, a new family of asiloid flies (Diptera: Brachycera: Asiloidea), based on Ocoa chilensis gen. and sp.n. from Chile, South America. Systematic Entomology 28: 417-431.
- Yucel, A, (1988) Investigation on determining flour beetles and their damages in meal factories and mills in southeastern Anatolia, Turkey. Bitki Koruma Bulteni 28: 57-78.
- Zaitsev, Z.F. (1972a) Some new species of beeflies (Diptera, Bombyliidae) from middle Asia and Kazakchstan. Zoologicheskii Zhurnal 51: 455-58.
- Zaitsev, Z.F. (1972b) On the fauna of beeflies (Diptera, Bombyliidae) of Mongolia, I. In Insects of Mongolia No. 1. Leningrad.
- Zetterstedt, J.W. (1844) Diptera Scandinaviae . vol. 3, pp. 895-1012. Officina Lundbergiana, Lundae [=Lund.].
- Zetterstedt, J.W. (1859) Diptera Scandinaviae . vol. 13, pp. xvi+ 4943-6190. Officina Lundbergiana, Lundae [=Lund.].
- Zuraska, I. (1975) Zarys morfologii I biologii oraz znaczenie gospodarcze tanecznicy okiennej, Omphrale fenestralis (L.) Ochrona Roslin 2: 20-22.
- Zuranska, I. (1979) Morfologia poczwarki Omphrale fenestralis (L.) (Diptera, Omphralidae) [The morphology of the pupa of Omphrale fenestralis (L.) (Diptera, Omphralidae)]. Przeglad Zoologiczny 13: 53-56.
- Zuska, J. (1987) Check list of Czechoslovak insects. 2. (Diptera) Scenopinidae. Acta Faunistica Entomologica Musei Nationalis Prague 18: 119.
Caenotus Cole 1923a: 14
Acaenotus Nagatomi & Yanagida in Nagatomi, Liu & Yanagida 1994: 142
Alloxytropus Bezzi 1925b: 186
Jackhallia Nagatomi & Liu 1994: 166
Proratites Grimaldi & Cumming 1999: 29
Belosta D.E. Hardy 1944a: 37
Brevitrichia D.E. Hardy 1944a: 32
Caenoneura Kröber 1923: 75
Heteromphrale Kröber 1937: 221
Irwiniana Kelsey 1971b: 279
Metatrichia Coquillett 1900: 500
Neopseudatrichia Kelsey 1969: 299
Paramonova Kelsey 1970b: 125
Paratrichia Kelsey 1969: 320
Paratrichia Kelsey 1969: 320
Paratrichia Kelsey 1969: 320
Prepseudatrichia Kelsey 1969: 286
Propebrevitrichia Kelsey 1969: 281
Pseudatrichia Osten Sacken 1877: 276
[=Atrichia Loew, 1866: 42]
Pseudomphrale Kröber 1913: 180
Riekiella Paramonov 1955: 652
Scenopinus Latreille 1802: 463
[=Archiscenopinus Enderlein 1914: 25]
[=Astoma Lioy 1864: 762]
[=Atrichia Schrank 1803: 54]
[=Cona Schellenberg 1803: 66]
[=Hypseleura Meigen 1803: 273]
[=Lepidomphrale Kröber 1913: 182]
[=Lucidomphrale Kröber 1937: 222]
[=Omphrale Meigen, 1800: 29 (suppressed)]
[=Omphralosoma Kröber 1937: 222]
[=Paromphrale Kröber 1937: 222]
[=Scenopoeus Agassiz, 1846: 333] [unjustified emendation]
[=Scaenopius Dalla Torre, 1878: 161] [misspelling]
[=Scenopinula Paramonov 1955: 651]
Seguyia Kelsey 1980: 323
[=Seguyella Kelsey 1969: 294]
Stenomphrale Kröber 1937: 220
SUBFAMILY incertae sedis
Cyrtosathe Winterton & Metz 2005 : 4
Cyrtosathe Winterton & Metz 2005 : 4
(modified from Kelsey 1969, etc., Yeates 1992, Nagatomi et al., 1994)
| 1a. Wing vein M2 absent (Scenopininae) | 3 |
| 1b. Wing vein M2 present | 2 |
| 2a. Costal vein extending around wing; sensory area on tergite 2 as two hemispherical regions of short setae; gonocoxal apodemes short; aedeagus straight; female tergite 8 with short setae (Caenotinae) [Nearctic] | Caenotus |
| 2b. Costal vein ending just beyond vein R5; sensory area on tergite 2 as two hemispherical regions of short setae; gonocoxal apodemes elongate; aedeagus recurved, folded dorsally upon itself; female tergite 8 dorsally covered with greatly elongate setae (subfamily incertae sedis) [Afrotropical - Namibia] | Cyrtosathe |
| 2c. Costal vein ending at vein R5; sensory area on tergite 2 as triangular region of setae with truncate apices; gonocoxal apodemes greatly elongate; aedeagus straight; female tergite 8 with short setae (Proratinae) | 20 |
| 3a. Cell R5 open to the tip of wing | 4 |
| 3b. Cell R5 closed and petiolate | 9 |
| 4a. Vein M1+2 fading out before edge of wing | 5 |
| 4b. Vein M1+2 reaching edge of wing | 6 |
| 5a. Vein R4 branching before middle of cell R5 [Australian] | Scenopinus (part, S. pallidipennis) |
| 5b. Vein R4 branching at or beyond middle of cell R5 [Australian] | Riekiella (part) |
| 6a. Vein CuA1 normal, reaching edge of wing | 7 |
| 6b. Vein CuA1 fading out beyond the crossvein m-cu [Afrotropical] | Seguyia |
| 7a. Head longer than high; long slim flies; often glossy black (resembling Pseudatrichia) [Afrotropical] | Prepseudatrichia |
| Scenopinus ........... 8 | |
| 8a. Large robust flies; antennae long, slender; vein R4 branching from R5 before middle of cell R5; male with 9th tergum 4 lobed | S. fenestralis-group |
| 8b. Medium sized flies; antennae variable; vein R4 branching from R5 near middle of cell R5; male with 9th tergum 2-lobed, open ventrally | S. albicinctus-group |
| 8c. Medium sized flies; antennae pear-shaped; vein R4 branching from R5 near middle of cell R5; male with basal portion of 9th tergum extending ventrally and with distal portion open; female with 8th sternum longer than tergum and 9th tergum usually with short spines | S. brevicornis-group |
| 8d. Medium to small flies; antennae short; vein R4 branching from R5 beyond middle of cell R5; male with 9th tergum 2-lobed, closed below | S. velutinus-group |
| 9a. Vein R5+M1+2 ending at tip of wing | 10 |
| 9b. Vein R5+M1+2 bending sharply to end in leading edge [Afrotropical] | Caenoneura |
| 10a. Long, slender black (usually glossy) flies | 11 |
| 10b. Brown, tan or gray flies | 13 |
| 11a. Head longer than high; body glabrous | 12 |
| 11b. Head higher than long, with mouthparts atrophied; body hairy [Neartic] | Belosta |
| 12a. Tip of cell R5 blunt; male with 9th tergum about as long as high, and as long as 10th tergum; female with 9th tergum and sternum subequal [Nearctic] | Pseudatrichia |
| 12b. Tip of cell R5 acute; male with 9th tergum much higher than long, and shorter than 10th tergum; female with 10th tergum with thick spines [Australian] | Neopseudatrichia |
| 13a. Large robust bodied flies | 14 |
| 13b. Smaller flies | 15 |
| 14a. Very large flies with broad, blunt abdomens in both sexes; frontal area swollen so that antennae project from middle of head; hairs often flat, scale like [Cosmopolitan] | Metatrichia |
| 14b. Smaller flies with blunt abdomens; frons not swollen; male genitalia with flange-like lobes on inside of 9th tergum [Australian] | Paratrichia |
| 15a. Pollinose scales [pruinescence?] on thorax | 16 |
| 15b. Black shiny flies wthout scales | 19 |
| 16a. Wing with vein R4 branching from cell R5 near the base | 17 |
| 16b. Wing with vein R4 branching from cell R5 near the middle | 18 |
| 17a. Male with two long aedeagal spines; female with 8th sternum excavated on distal margin, 9th tergum often with row of stiff spines on posterior margin [Nearctic, Neotropical] | Brevitrichia |
| 17b. Male with short aedeagal parameres; female with long pointed 8th sternum, 9th tergum with stiff spines [Neotropical] | Heteromphrale |
| 18a. Male with 9th tergum as two flaps covering genitalia; female with distinctive spines on 9th tergum [Afrotropical] | Propebrevitrichia |
| 18b. Male with 9th tergum as two lobes, short; female with reduced spines on 9th tergum, 8th sternum longer than tergum, pointed [Australian] | Riekiella (part) |
| 18c. Male with 9th tergum as four long lobes, elongate; female with 8th segment elongate, blunt, tergum and sternum subequal [Australian] | Paramonova |
| 18d. Male with 9th tergum as two [?] short lobes; female with reduced spines on 9th tergum, 8th sternum longer than tergum, 3-lobed apically [Neotropical] | Irwiniana |
| 19a. Antennae pointed; vein R4 branching from R5 at distal third of cell R5 [Afrotropical] | Stenomphrale |
| 19b. Antennae blunt; vein R4 branching from R5 before middle of cell R5 [Palearctic] | Pseudomphrale |
| 20a. Antennal flagellum gradually tapered apically, triangular or lancet shaped; apical style small and inconspicuous; probosis largely sclerotised, usually longer than face; mesonotum and scutellum with bristles; thickening of costa eding at or just beyond apex of R5 | 22 |
| 20b. Antennal flagellum not as above; probosis fleshy and shorter than face; mesonotum without bristles; scutellum with or without bristles | 21 |
| 21a. Antennal flagellum abruptly narrower at apical portion, with a tuft of hairs at apex, without apical style; thickening of costa ending at or just beyond apex of R4; scutellum without bristles; abdomen largely white or pale yellow [Nearctic] | Caenotoides |
| 21b. Antennal flagellum gradually narrowed apically, with thick apical style that is wider than apex of preceding segment; thickening of costa ending at or just beyond apex of R5; scutellum with a pair of marginal bristles; abdomen dark brown or black [Nearctic] | Acaenotus |
| 22a. Vein M2 arising from M1; female occiput not forming postocular rim | 23 |
| 22b. Vein M2 arising from discal cell; female with postocular rim [Neotropical] | Jackhallia |
| 23a. [Nearctic] (still need a good character!) | Prorates |
| 23b. [Afrotropical] (still need a good character!) | Alloxytropus |
Entomology Sub-Menu
Plant Pest Diagnostics Center
CDFA Collections
General Info/Links
Contact Us
3294 Meadowview Road
Sacramento, CA 95832
Phone: 916-262-1100
Fax: (916) 262-1190